
You
are here: Home
> Introduction
> Nitric Oxide
> Regulation of Nitric Oxide production
|
|
Regulation of NO production
Nitric oxide is an important signalling molecule but is also highly reactive and highly diffusable. It is therefore important that there is strict control and regulation of nitric oxide production. The synthesis of NO within cells can be regulated in a number of ways such as the cellular distribution of NOS, changes in NOS gene expression, enzymatic activation by phosphorylation and the presence of cellular inhibitors NOS activity.
Intracellular distribution of NOS
One of the most important ways in which NO is regulated is through strict control over the location of NO production.
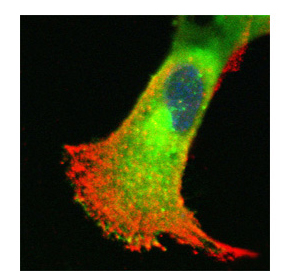 The NOS isoforms can be targeted to different regions of the cell, where NO will be produced in close contact with its target proteins. The image on the left shows the distribution of iNOS (shown in green) and eNOS (shown in red) in a trophoblast cell. The nucleus is shown in blue. Co-localisation between iNOS and eNOS will show up as a yellow colour.
The NOS isoforms can be targeted to different regions of the cell, where NO will be produced in close contact with its target proteins. The image on the left shows the distribution of iNOS (shown in green) and eNOS (shown in red) in a trophoblast cell. The nucleus is shown in blue. Co-localisation between iNOS and eNOS will show up as a yellow colour.
It can be seen that iNOS and eNOS show quite distinct distribution in the cell, with very little yellow colour suggesting very little overlap in their cellular distribution. Although both iNOS and eNOS produce NO it is likely that their different cellular distribution will lead to NO interacting with different targets and therefore having different effects.
It is known that eNOS can be targeted to the plasma membrane, the Golgi complex and the caveolae. In the latter case interaction between eNOS and caveolin-1 is known to inhibit eNOS activity.
The distribution of NOS isoforms in the cell is likely to be an important mechanism for regulating where and when NO is produced. As such the transport of NOS within the cell, the signals that trigger NOS redistribution and the proteins that NOS interacts with are important ongoing areas of investigation.
Activation of NOS activity
One of the main activators of NOS enzyme activity is changes in cellular calcium levels. The constitutive isoforms of NOS, eNOS and nNOS, show increased activity following increases in calcium, and therefore calmodulin, in the cell. In addition both eNOS and nNOS are known to show further increases in activity following phosphorylation. There are at least 5 regulating phosphorylation sites on eNOS and these can be regulated by a number of different kinases and phosphatases, with the protein kinase Akt being one of the best understood regulators of eNOS activity. It is also known that nNOS has both activating and inhibiting phosphorylation sites, but much less is known about the post-translational regulation of iNOS. There is some suggestion that iNOS activity can be regulated by Src mediated tyrosine phosphorylation, but it appears that iNOS activity is mostly regulated at the transcriptional level and through its intracelular distribution.
Endogenous inhibitors of NOS
The synthesis of NO can also be inhibited by two methylated analogues of arginine, asymmetric dimethylarginine (ADMA) and mono methylarginine (L-NMMA). These inhibitors are endogenously produced competitive inhibitors of NOS formed by the post-translational methylation of arginine residues in proteins and liberated upon their hydrolysis.
Free ADMA is found in plasma and urine, while the intracellular concentration of ADMA is about 5 times higher than the extracellular concentration. We have shown that circulating ADMA falls during normal pregnancy but is elevated early in pregnancies complicated by pre-eclampsia. There is also evidence that elevated levels of ADMA are important in a number of other conditions including diabetes, atherosclerosis and renal failure. Interestingly, the administration of NOS inhibitors to pregnant rats results in features similar to pre-eclampsia, including a reduction in placental size and utero-placental blood flow, both of which can be reversed by the administration of L-arginine, suggesting a specific NO-mediated effect. Within cells the concentration of ADMA and L-NMMA, but not symmetric dimethylarginine (SDMA), is regulated by the activity of the cytoplasmic enzyme dimethylarginine dimethylaminohydrolase (DDAH). Inhibition of DDAH activity leads to elevated ADMA in culture and inhibits endothelium-dependent relaxation. These experiments demonstrate that DDAH is basally active and that inhibition of DDAH leads to local accumulation of ADMA which reaches concentration sufficient to inhibit NOS.
Website by Phil Dash